- Visibility 295 Views
- Downloads 55 Downloads
- Permissions
- DOI 10.18231/j.ijpca.2024.017
-
CrossMark

Extracellular vesicles the delivery of drugs through genetic engineering
- Author Details:
-
 Joshi Ankur *
Joshi Ankur *
-
Soni Priyanka
-
Malviya Neelesh
-
Malviya Sapna
-
Kharia Anil
Abstract
Researchers from a variety of disciplines are investigating the use of extracellular vesicles (EVs) as delivery vehicles for pharmaceutical solutions. In order to make the most of the therapeutic potential of EVs, various loading strategies have been devised. Despite the widespread application of exogenous methods, endogenous approaches are becoming increasingly preferred instead. Medicinal macro molecules such as proteins and nucleic acids can be loaded with relative ease using this technology that manipulates parental cells through genetic engineering. We examine the most effective ways for EV loading and demonstrate why endogenous is superior. Extracellular vesicles (EVs), which are able to transport pharmaceuticals, have the potential to tackle the problems of biologic therapeutic stability and off-target undesirable effects. In addition, we investigate the most recent findings and applications of this innovative method in order to shed light on the numerous potential therapeutic possibilities in the field of EV-based treatments. The possibility of endogenous loading of EVs with big biological medicines has been brought to light by recent research. In order to optimize the loading of biologic drugs in extracellular vesicles (EVs), researchers have been able to apply cargo selection and loading pathway molecules thanks to advancements in EV biogenesis. It has been established that endogenous EV loading is successful in preclinical in vivo trials, which demonstrates its promise in a variety of therapeutic situations.
EV Nanoprobes Have the Ability to Supply Biomolecules
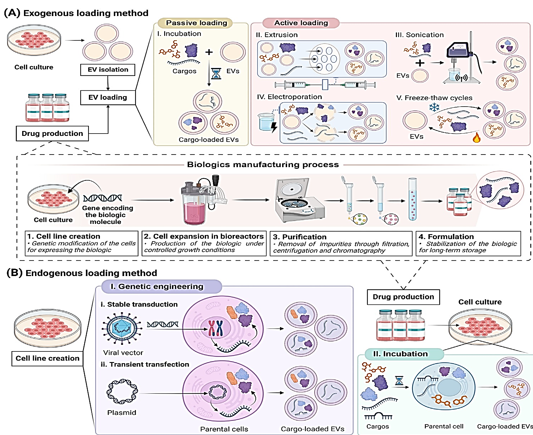
As stated in the Glossary, biologics have accounted for thirty percent of all FDA-approved drugs since 2014. By the year 2023, five of the fifty-five new medications were biosimilars. 15 biosimilars have been authorized by the FDA as of this moment. Out of the 39 new medications that were approved by the EMA in 2023, eight of them were biosimilars. The FDA has given its approval to eighty biosimilars since 2006. The approvals demonstrate an increase in the use of biologic treatments. The stability concerns of these drugs, which are caused by their susceptibility to fluctuations in temperature, solvent, and pH, as well as on-target off-tissue side effects, need to be addressed.[1] Liposomes and other artificial nanoparticles have been used for transporting biologic medications for a long time.[2], [3] Recently, however, there has been a resurgence in interest in nanomedicine as a more effective drug delivery vehicle for the treatment of a wide variety of disorders.[4], [5] This is because to the exceptional targeting capabilities, improved safety profile, and high biocompatibility of extracellular vesicles. It is possible that extracellular vesicles are more effective at transporting medicine than nanoparticles.[6] The fact that they are able to genetically change parental cells gives them the potential to manufacture and integrate therapeutic biomolecules into EVs using their own machinery. This is only one of the numerous benefits that they have over conventional drug delivery methods. When compared to external loading, endogenous loading can make the process of manufacturing, purifying, and storing biological drugs more straightforward[7] (see [Figure 1] BI). There is a possibility that EVs will enhance the therapeutic efficacy of new biologic medicines and help offset some of their disadvantages.In this research, we discuss the various endogenous ways for creating and loading biological materials into extracellular vesicles (EVs), as well as the existing strategies for loading extracellular vesicles (EVs). For the purpose of shedding light on the numerous EV-based therapies, we will examine the most prevalent methods to maximize EV cargo loading and the therapeutic applications of these methods.
Loading Instructions for Endogenous Extracellular Vesicles
Extracellular vesicles can be loaded either from the outside or from the inside. The loading of synthetic nanoparticles into extracellular vesicles (EVs) has been accomplished by the use of exogenous methods, as demonstrated in Figure 1A. However, these approaches are detrimental to EVs because they modify the structure and membrane stability of the EVs ([Table 1]). It is possible for targeting and immunocompatibility to be compromised when necessary membrane proteins are broken down.[8] Extracellular vesicles and the payloads they carry could be damaged by forces from the outside. [9], [10] Therefore, endogenous methodologies, which are not harmful to extracellular vesicles (EVs) and do not involve their destruction, have been widely used in research ([Table 1]).
At the time of EV biogenesis, the endogenous system, also known as the pre-encapsulation mechanism, loads cargo. It is possible to load cargo through endogenous incubation or genetic engineering. The incubation process comprises loading extracellular vectors (EVs) onto cargo and then adding it to the cell culture medium in a concentration gradient, as shown in [Figure 1] BII. Because of this, parental cells are able to integrate cargo into their cytoplasm." Drugs that are loaded into incubation must not cause harm to the cells of the parents.[11] The purpose of genetic engineering is to boost the expression of therapeutic biomolecules by modifying the DNA of the parent cells. The target molecule is synthesized by the body, and then it is encased in energetic vehicles (EVs),which is a novel approach.[12] When it comes to loading extracellular vectors (EVs) with therapeutic macromolecules, genetic engineering is a more integrated and straightforward way than incubation and exogenous procedures. In this study, the term "endogenous method" is used exclusively to refer to the process of genetic engineering of parental cells. This method is the most efficient way to increase the loading efficiency of proteins and nucleic acids.[13] Scientists loaded biologic pharmaceuticals onto EVs by the use of stable transduction and transient transfection of parental cells. Lentiviruses are utilized in the process of transduction in order to introduce the therapeutic payload gene into the DNA of the cell.[14], [15] This can be seen in [Figure 1] Bi. As demonstrated in Figure 1Bii, the process of transfection involves the introduction of therapeutic oligonucleotides or plasmids that can express the therapeutic gene into parental cells. Use is made of a number of different therapeutic oligonucleotides. mRNAs, circRNAs, long noncoding RNAs, and siRNAs are some examples. Depending on the gene that is being silenced, these RNAs can either temporarily or permanently silence the epigenetic expression of the gene. [16] EVs can also be produced by transformed cells using the chemical that is produced by the overexpressed gene. Because of this, extracellular vesicles are able to transport growth factors and cytokines.[17], [18], [19], [20], [21] Numerous specialists have demonstrated that genetically produced extracellular vesicles are able to maintain their physicochemical qualities more effectively than exogenous loading methods ([Table 1]). For the purpose of treating lung injury, Salazar-Puerta and colleagues modified dermal fibroblasts to secrete extracellular vesicles (EVs) that expressed the anti-inflammatory cytokines IL-4 and IL-10. The genetic engineering loading did not cause any damage to the EV lipid membrane, according to the findings of these researchers.[21] Lu and colleagues developed a therapy for skin lesions that is based on genetic engineering. A cell line with robust NF-κB siRNA secretion was created by researchers by the use of lentiviral transduction of mesenchymal stem cells (MSCs) derived from adipose tissue. At the same time, non-modified cells served as controls. The size and expression of membrane indicators were identical in exosomes from control cells loaded with siRNA and those without.[22]
|
S. No. |
Aspect |
Exogenous Loading |
Genetic Engineering |
|
1. |
Cargo type |
Typically employed for the loading of microRNAs and tiny synthetic medicines. |
Capable of loading larger proteins and nucleic acids. |
|
2. |
Examples of Molecules loaded |
It includes several small compounds with molecular weights less than 1 Kilodalton: 1. Resveratrol (228.25 Dalton) 2. Hydroxychloroquine (335.87 Dalton) 3. Curcumin (368.38 Dalton) 4. Doxorubicin (544.82 Dalton) 5. Paclitaxel (853.91 Dalton). |
It includes several large proteins with molecular weights more than 10 Kilodalton: 1. IL-12 (75 kilodalton) 2. IL-10 (18.6 kilodalton) 3. GDNF (23 kilodalton) 4. BDNF (27 kilodalton) 5. PEDF (50 kilodalton) |
|
3. |
Efficient Loading of Nucleic Acids |
Larger molecules (>1 Kilobyte) are less efficient than smaller nucleic acids, such as siRNAs and miRNAs, which include 18-25 nucleotides. |
Both short (<1 Kilobyte) and large (>1 Kilobyte) nucleic acid molecules can be efficiently processed. |
|
4. |
Sustaining EV Integrity |
Might lead to EV aggregation, changes in size, and changes in membrane properties. |
Preserves the integrity of EVs while preserving their physicochemical features. |
|
5. |
Challenges in Optimization |
Accurate control of experimental variables such as shear force, electric field voltage, freeze-thaw cycles, and surfactant concentration is necessary to minimise damage to the extracellular vesicles (EVs). |
It is necessary to have strong control over the variables that impact the production of extracellular vesicles (EVs), such as production time and cell confluence, in order to ensure uniformity across batches. |
|
6. |
Impact on extracellular vesicle Sample Quality |
Entails iterative processes of isolating and refining, which might deteriorate extracellular vesicles (EVs) and diminish the quality of the sample. |
Maintains the integrity of EV samples without the use of further purifying procedures. |
|
7. |
Requirements for Essential Equipment |
Specialised equipment is required |
There is no need for any further equipment. |
Enhancing EV Biologic Drug Loading Via Engineering
It is necessary to have an understanding of the complex cargo integration processes in order to load biologics into endogenous vehicles. The cargo sorting process for EV production involves a number of molecules and is subject to stringent supervision. [23] Recent advancements in molecular biology, mass spectrometry, and next-generation sequencing have led to the discovery of molecules that are responsible for the selection and loading of extracellular vesicles cargo. [24] Having this insight has resulted in the development of novel ways for loading EV biologics. [25] These forward-thinking technologies include molecular sorting modules, which are responsible for adding EV payload. Scientists have the ability to genetically modify parent cells using MSM in order to increase the expression of biologic drugs. The conversion of biologics into EVs occurs naturally during biogenesis. [26]
.
Methods developed recently to improve EV protein medication loading
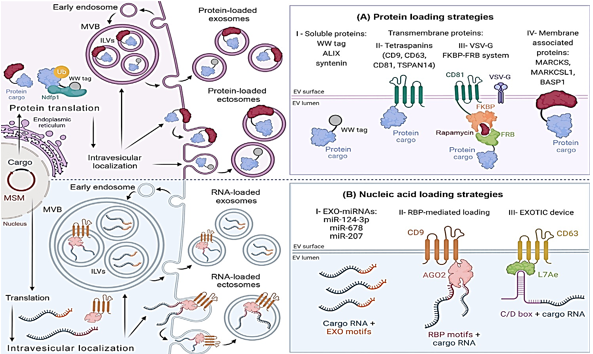
Improvements in protein cargo loading have been made possible by a variety of methods that imitate EV protein loading systems. By recognizing and encapsulating ubiquitin-tagged proteins into extracellular vesicles, the late-domain route, also known as the L-domain pathway, is responsible. In order to carry out this plan, the NEDD4 ubiquitin ligase family engages in a conversation with NDFIP1. According to the findings of the research, WW tags are attached to target proteins by the L-domain motifs of NDFIP1. Protein motifs known as WW tags consist of two tryptophan (W) residues that are next to one another. NEDD4 is able to ubiquitinate and integrate into exomes with greater ease when the cargo protein in question has a WW tag.[27] NDFIP1 is responsible for the packaging of WWTR1, also known as TAZ, into exosomes, as Cheng et al did. It is demonstrated in Figure 2AI that NDFIP1 is the only one that can identify TAZ with its WW domain intact, in contrast to cTAZ, which is a shortened TAZ that does not contain it. Intriguingly, this demonstrates that NDFIP1 only interacts with TAZ and not with cTAZ. Tetraspanin-rich microdomains are responsible for loading extracellular vesicles with proteins. In order to develop an effective medium for the transport of extracellular vectors (EVs), Zuppone et al. genetically incorporated an RFP reporter protein onto CD9, CD63, and CD81 tetraspanins. CD9-RFP was selected as the most effective recombinant protein due to its ability to be delivered to recipient cells and extensively sorted onto extracellular vectors (EVs). Using an EV MSM that has been folded and sorted can help prevent lysosomal degradation[28] There is also a significant load of tetraspanin TSPAN14. In contrast to CD63 and other EV biogenesis proteins such as ALIX and syntenin, which can only load between 40 and 70 EGFP/vesicles, it was able to load 150 EGFP/vesicles.[29], [30] The efficiency with which cargo is loaded is impacted by cargo attributes, which makes it challenging to select the appropriate MSM. These procedures are straightforward to adopt and make advantage of biological mechanisms to prevent injury to parent cells. Additionally, the vesicular stomatitis virus G is capable of loading EVs. Multiple studies have demonstrated that VSVG is capable of loading extracellular vesicles with a wide variety of chemicals. [31] Fusing EV membranes is one of the ways that VSVG helps to increase cell internalization and cargo release.[32] The endosomal escape pathway is a barrier to the EV-mediated intracellular administration of medicines. There have been several studies[33] that have shown that proteins that are given in this manner prevent the lysosomal breakdown of recipient cells.
VSVG-based synthetic EVs and rapamycin-regulated protein-protein controlled loading were both improved by Somiya and colleagues over the course of their research. FK506-binding protein (FKBP12) and the FRB domain are both complexed by rapamycin, which is a relatively tiny chemical. Somiya and his colleagues fused the target protein with FRB after first combining CD81 tetraspanin and FKBP from the previous step. Rapamycin improved the interaction between FKBP12 and FRB, which resulted in cargo being recruited to EVs ([Figure 2] AIII). The researchers believe that the viral origins of VSVG could potentially elicit immunological reactions, which would render it unsuitable for injection into living organisms. Therefore, proteins that are not viral are advised for use in treatment.[34] It has been demonstrated by Ilahibaks et al. that this method is capable of transporting intracellular protein in vivo, despite the constraints that were anticipated.[35] FRB/FKBP heterodimerization and syncytin-1, a naturally occurring human protein, were utilized by Bui et al. in the development of a method that is analogous to the one described above. Within the laboratory, this alternate fusogen is able to enhance both VSVG and cargo movement. [36] In order to enhance protein loading, therapeutic compounds can be combined with membrane-associated proteins that are created by electronic vehicles. Proteomic study led to the discovery of the MARCKS protein family, which includes MARCKS, MARKCSL1, and BASP1[37] (Figure 2AIV). This family has potential advantages. Research has shown that extracellular vesicles (EVs) are involved in a number of biological functions; however, it is not understood what role they play in the process of biogenesis. They change the dynamics of the actin cytoskeleton as well as the trafficking of vesicles [38].
The most recent techniques for enhancing the drug loading of EV nucleic acid
In addition, the intricate processes that control the loading of proteins and nucleic acids into extracellular vesicles are connected. Even while the composition of EVs is mostly determined by their mother cell, there are a variety of subtypes. [39] Exosomes have a greater number of microRNAs, according to a number of studies, while ectosomes have a genetic composition that is comparable to the transcriptome of the parent cell. MiR-124-3p, miR-678, and miR-207 have all been found to be expressed in exosomes, according to recent research. A higher level of miR-343 and miR-140-3p expression is seen in parental cells ([Figure 2] BI). Because of the unique patterns that miRNAs exhibit, miRNA loading is dependent on the sequence of DNA.[40], [41]
RBPs, like as hnRNPs, have the ability to load EVs with nucleic acids. Exosome miRNA trafficking is regulated by hnRNPA2B1, which is one of the hnRNPs that has received the most investigation. For the purpose of gene therapy, this chemical has the potential to load EXO-motifs miRNAs into the lumens of exosomes. [42] Research indicates that hnRNPA2B1 is responsible for modulating the packing of circRNA. [43] Searching for RBPs that target the inclusion of EV RNA is extremely important. Numerous investigations have uncovered more RBPs that are capable of accomplishing this. SYNCRIP, AGO2, VAP-A, YBX-1, FMR1, and HuR are some of the examples of these. Es-Haghi and colleagues derived hCD9.hAGO2 from their own personal experiences. Exosome biogenesis-related CD9 and RBP AGO2 are elements that are contained inside this vector (Figure 2BII). EVs that were transformed with the hCD9.hAGO2 vector were shown to contain higher quantities of miRNA compared to cells that overexpress miRNA, according to the findings of the study.[44] There is a greater abundance of the zipcode, which is a component of the 3′-untranslated region (UTR) of mRNA, in EVs.[45] Additionally, the 3'-untranslated region (UTR) of the mRNA of interest can be modified to include a sequence that is similar to a Zipcode. The loading of EV mRNA is improved by EXOTIC. Additionally, the rRNA packaging protein L7Ae and the C terminus of CD63 are utilized in this technique. A 3'-UTR C/D box is the mechanism by which it binds therapeutic mRNA (Figure 2BIII).
|
Therapeutic area |
Cargo |
EV source |
Disease |
Biological effect |
Ref. |
|
Cardiovascular diseases |
The microRNA miR-183-5p |
Bone marrow stromal cells |
Myocardial I/R injury |
By lowering FOXO1 levels, it decreases oxidative stress and cardiomyocyte death. |
|
|
The microRNA miR-302d-3p |
Mesenchymal stem cells |
Acute myocardial infarction |
Deactivates the NF-κB pathway, which decreases inflammation. |
||
|
The microRNA miR-342-3p |
Human stromal mesenchymal stem cells |
Deep vein thrombosis |
Reduces endothelial cell injury through lowering EDNRA expression |
||
|
The microRNA miR-19b-3p |
Macrophages |
Atherosclerosis |
Increases blood flow to tissues by activating genes that promote angiogenesis |
||
|
The microRNA miR-126 |
Human induced pluripotent stem cells |
Limb ischemia |
Stimulates dendritic cells to halt tumour growth |
||
|
Cancer |
Interleukin-12 |
Colon carcinoma cells |
Colon carcinoma |
Limit tumour growth by activating dendritic cells |
|
|
PEDF |
Human breast cancer cells |
Breast cancer |
Repolarizes macrophages from M2 to M1 |
||
|
DTA |
HeLa cells |
Cancer |
Causes tumour cells to die by blocking the production of new proteins. |
||
|
miR-1915-3p |
Nontumorigenic intestinal cells |
CRC |
By inhibiting the PFKFB3 and USP2 oncogenes, it makes oxaliplatin more sensitive. |
||
|
miR-506-3p |
Nontumorigenic intestinal cells |
CRC |
Downregulates GSTP1, which inhibits the development of cancer cells. |
||
|
miR-323a-3p |
Human CRC cells |
CRC |
Directs EGFR/TYMS-mediated cell death towards cancer cells |
||
|
miR-126-3p |
Human BMSCs |
Non-small cell lung cancer |
Inhibits the development of cancer cells by downregulating PTPN9 expression |
||
|
miR-142-5p |
Human NPC cells |
Radioresistance in NPC |
Enhances radiosensitivity by suppressing EGF/EGFR and HGF/c-MET |
||
|
miR-127-3p |
hucMSCs |
Horiocarcinoma |
Reduces EMT via decreasing ITGA6 levels |
||
|
miR-29b |
BMSCs |
Gastric cancer |
A decrease in fibronectin 1 expression inhibits cancer cell attachment to hematopoietic stem cells. |
||
|
circDIDO1 |
HEK 293T cells |
Gastric cancer progression |
Reduces cancer growth by boosting SOCS2 expression |
||
|
GSDMD-N mRNA |
HEK 293T cells |
Cancer immunotherapy |
Through pyroptosis, it instigates anti-tumor immunity. |
||
|
Nervous System Disorders |
BDNF |
MSCs |
Cerebral ischemia |
Protects neurons and reduces inflammation |
|
|
DNAJB6b chaperone |
NSCs |
Huntington disease |
Decreases the aggregation of mutant HTT via blocking the aggregation of PolyQ. |
||
|
TPP1 enzyme |
Macrophages |
Batten disease |
Promotes autophagy to break down aggregates of lipofuscin |
||
|
miR-410 |
BMSCs |
Hypoxic/ischemic brain damage |
Attenuates neuronal apoptosis by inactivating the WNT pathway through HDAC4 targeting |
||
|
miR-130a-3p |
ADSCs |
Diabetic peripheral neuropathy |
Reducing DNMT1 levels promotes Schwann cell proliferation |
||
|
circDYM |
HEK 293T cells |
Major depressive disorder |
Reducing levels of genes that are downstream targets of TAF1 reduces neuroinflammation. |
||
|
Respiratory diseases |
IL-4 and IL-10 |
Dermal fibroblasts |
Acute lung injury |
Lessens tissue damage by blocking the release of pro-inflammatory cytokines. |
|
|
miR-26a-5p |
hucMSCs |
Silica-induced lung fibrosis |
Reduces EMT caused by silica by blocking the ADAM17/NOTCH signalling route. |
||
|
Renal diseases |
IL-10 |
Macrophages |
Ischemic acute kidney injury |
Reduces mTOR signalling to maintain mitochondria in TECs |
|
|
ACE2 |
hucMSCs |
Renal I/R injury |
Prevents kidney damage caused by ischemia-reperfusion by triggering the NRF2/HO-1 response. |
||
|
GDNF |
ADMSCs |
Tubulointerstitial fibrosis |
Alleviates peritubular capillary loss through SIRT1/eNOS pathway activation |
||
|
miR-186-5p |
Human MSCs |
Renal fibrosis |
Prevents EMT through obstructing the TGF-β1/Smad5 pathway. |
||
|
miR-26a |
HEK 293 cells |
Renal tubulointersticial fibrosis |
Lessens CTGF expression through SMAD3 activation inhibition |
||
|
Musculoskeletal diseases |
miR-92a-5p |
Prostate cancer cells |
Osteoblastic bone disease |
Reduces MAPK1 and FOXO1 expression, which increases osteoclast function. |
|
|
miR-140-5p |
hUSCs |
Knee osteoarthritis |
Improves secretion of the extracellular matrix by reducing VEGFA |
||
|
miR-370-3p |
Macrophages |
Osteoarthritis |
Mitigates chondrocyte inflammatory responses through SOX11 inhibition |
||
|
circPARD3B |
MSCs |
Osteoarthritis |
Regulates angiogenesis via decreasing VEGF expression |
||
|
circ_0050205 |
Mouse BMSCs |
Intervertebral disc degeneration |
limits ECM degradation and nucleus pulposus cell death through GPX4 upregulation. |
||
|
FOXF1 mRNA |
Human NP cells |
Intervertebral disc degeneration |
Transforms NP cells into a healthy phenotype |
||
|
Bmp2 mRNA |
293T cells |
Bone regeneration |
Encourages the process of bone repair by boosting osteogenesis |
||
|
DMD exon 53-targeting RNP |
HEK 293T cells |
DMD |
Edits exon 53 to enhance dystrophin expression |
||
|
Dermatological disorders |
NF-κB siRNA |
ADMSCs |
Skin Lesion |
Downregulates inflammatory cytokine production through blocking the NF-κB pathway. |
|
|
miR146a |
PMSCs |
Diabetic Wound Healing |
Minimises inflammatory response by preventing IRAK1 expression |
||
|
HOTAIR |
MSCs |
Chronic Wounds |
Upregulates VEFG, which promotes angiogenesis and stimulates wound healing. |
||
|
lncRNA H19 |
ADSCs |
Skin Photoaging |
Upregulating SIRT1 prevents DNA damage and the generation of MMPs and ROS |
||
|
COL1A1 mRNA |
Dermal fibroblasts |
Skin Photoaging |
Encourages the production of collagen protein grafts and decreases the onset of wrinkles. |
||
|
Inflammatory diseases |
IL-27 |
MSCs |
Inflammatory bowel disease |
Regulates T cell differentiation to inhibit inflammation. |
|
|
miR-126 |
293T/17 cells |
Periodontitis |
Facilitates the polarisation of macrophages to fight inflammation |
||
|
Infections and viral diseases |
ZPAMt mRNA |
HEK 293T cells |
HIV-1 infection |
Reduces HIV-1 expression by epigenetic mechanisms including DNA methylation |
|
|
Metabolic disorders |
LDLR mRNA |
HEK 293T cells |
Familial hypercholesterolemia |
Lowers the risk of atherosclerosis and liver steatosis via boosting hepatocyte LDLR expression |
In some cases, exoTIC devices may contain CX43 protein. Therapeutic messenger RNA is transported to the cytoplasm of the target cell by CX43.[85], [83]
Treatment using EVs loaded with GMOs.
The ability of genetically engineered extracellular vectors (EVs) to transfer biomolecules into specific cells for therapeutic purposes has been demonstrated by a number of research.[52] There is considerable potential for the in vivo translation of in vitro discoveries.[63], [79] The preclinical models of genetically designed loaded EVs that carry biologics for the treatment of cancer, cardiovascular disease, and disorders of the nervous system show promise. [46], [53], [61] As shown in. [Table 2]
The selection of an EV cell source is an essential step in the creation of EV pharmaceuticals. Evolved cells (EVs) usually maintain the traits and biological functions of their parent cells.[86] EVs produced from MSCs increase the communication between immune cells and the development of blood vessels. However, EVs that are produced from immune cells carry MHC molecules, which have the ability to either stimulate or repress immunological responses. EVs that are produced from tumors are able to target particular body regions and encourage the development of tumors.[87], [88], [89] The biological features of each and every EV-producing cell need to be assessed in order to provide a safe and effective treatment.
Genetically Modified Extracellular Vesicles for Various Diseases
Transportation vehicles that Have been genetically modified for the purpose of treating cardiovascular disease
Cardiovascular diseases pose a threat to the health of people all over the world and call for the development of novel treatments. Recent research has investigated the possibility of using genetically modified loaded extracellular vesicles (EVs) as a treatment for cardiovascular disorders. Contrary to conventional medical practices, therapies based on nucleic acid have the potential to cure or reduce symptoms. This can be accomplished through the regulation of gene expression as well as intracellular signaling pathways.[1] 2. An investigation into the protective effects of miR-183-5p-overexpressing MSC-derived exosomes against myocardial ischemia/reperfusion was carried out by Mao et al. The microRNA MiR-183-5p was able to permeate cardiomyocytes, hence reducing oxidative stress and mortality in cardiomyocytes that had been separated or decreased. [46] This resulted in a considerable improvement in cardiac function. A similar experiment was carried out by Liu and colleagues in order to develop an AMI treatment. After that, the researchers investigated the inflammatory environment of AMI as well as the molecular mechanisms involved in heart remodeling. AMI mice exhibited a decrease in the expression of miR-302d-3p. It was discovered in the study that EVs loaded with miR-302d-3p improved the health of the heart in AMI mice. Significant reductions were observed in the infarcted area, myocardial fibrosis, inflammation, apoptosis, and cardiac dysfunction. MiR-302d-3p was loaded into MSC-derived extracellular vesicles using their endogenous mechanism.[47] There have been recent developments that point to heart-specific nucleic acid therapies.
EVs that fight cancer have been genetically engineered
Both treatment resistance and metastasis are factors that influence cancer mortality rates internationally. A number of studies have demonstrated that extracellular vesicles are capable of transporting messenger RNAs, microRNAs, circRNAs, and proteins. According to the findings of these investigations.[59], [19], [53], [60] EVs have the potential to be an effective and selective cancer treatment.[Figure 2] is the reference.
The synthesis of the diphtheria toxin (DTA) catalytic A domain was increased through the use of genetic modification of parental cells by Dancourt et al. Because it prevents the production of proteins, this toxin is lethal to cancer cells. The cell cannot be reached by DTA on its own, even when it is present at large concentrations in the extracellular media. On the other hand, DTA in the cytoplasm is lethal to cells. EVs were shown to be capable of transporting DTA into the cytoplasm of recipient cells by the authors. The creation of proteins was inhibited, and cancer cells were eliminated as a result. [52] In order to investigate the efficacy of DTA for tumor ablation, researchers have utilized other carriers such as viruses and liposomes.[90] On the other hand, problems with manufacturing, formulation, and immunogenicity have held down the development process. It is easier to create and produce extracellular vesicles DTAs when endogenous loading is used. Since EVs transport poisons in a more natural manner than liposomes or viruses, they are less likely to be rejected. [52] Xiao et al. conducted a separate experiment in which they found that genetically altered loaded EVs were effective in treating colorectal cancer that was resistant to oxaliplatin. While the expression of miR-1915-3p was much higher in oxaliplatin-resistant cell lines, it was significantly lower in non-tumorigenic intestinal cells. The therapeutic miRNA was transmitted to a colorectal cancer cell line by first being overexpressed in a nontumorigenic intestinal cell line and then being transferred to the colorectal cancer cell line for use. MicroRNA-1915-3p was found to reduce drug-resistant cancer-causing genes in mice xenograft models, which resulted in an increase in oxaliplatin sensitivity. [53] Pang et al. conducted research on 5-fluorouracil (5-FU) chemotherapy with the goal of enhancing the treatment of colorectal cancer. They developed colorectal cancer tumor cell lines that overexpress miR-323a-3p for the reason that tumor-derived exosomes are attracted to the cells from which they originated. Exosomes that carry miR-323a-3p preferentially target tumors, hence lowering growth and enhancing 5-FU tumor killing. [55] Finding a safe cancer treatment that decreases side effects from chemotherapy and radiotherapy is another important challenge. The results verified a commonly held opinion that has been generally recognized for a long time. For the purpose of addressing this matter, Guo et al. investigated the impact that exosomes carrying circDIDO1 have on the progression of gastric cancer. The therapeutic circRNA was successfully carried to the cancer cells in the stomach by the altered exosomes. As a result, the circRNA was able to absorb miR-1307-3p, which led to a rise in SOCS2 expression.
The cytokine-induced signal transduction that is inhibited by SOCS2 results in a reduction in the proliferation, migration, and invasion of tumor cells. Exosomes loaded with circDIDO1 were administered to mice, and histological examinations revealed no abnormalities or lesions in any of the vital organs, including the kidney, spleen, heart, liver, or lungs. This provides evidence that exosomes have the potential to be successful therapies for stomach cancer.[59] EVs that have been engineered can also transport mRNA, miRNAs, and circRNAs for the therapy of cancer. mRNA, in contrast to other nucleic acids, communicates genetic information from DNA to ribosomes, which are responsible for the production of proteins.[16] However, there is a substantial possibility that EV-delivered mRNA might be used to precisely regulate the production of proteins in particular cells. Researchers Xing et al. used extracellular vectors (EVs) to deliver GSDMD-N mRNA to cancer cells. The GSDMD-N protein was produced as a result of the translation of the mRNA payload in the target cells. Because of pyroptosis, the tumor expanded at a slower rate. GSDMD-N administration through endogenous loading is associated with a number of disadvantages. It should be noted that parental cells may go through pyroptosis prior to the translation of mRNA and the release of EV. In order to find a solution to this issue, puromycin was utilized to inhibit the translation of parental cell mRNA. By using this approach, cells were preserved, and EVs were able to encapsulate mRNA. EV sorting is prevented when mRNA is translated correctly in parent cells because it binds to ribosomes and prevents it from moving around. The increased incidence of noncoding RNA in EVs can be explained by this strategy. Innovative technique developed by Xing and colleagues for encapsulating messenger RNA (mRNA) in extracellular vesicles(EVs) contributed to the advancement of scientific knowledge, [60] GM-loaded EVs for the treatment of disorders of the nervous system
EVs are fascinating as potential therapies for neurological problems
because of their immunological privilege and their ability to traverse the blood-brain barrier.[91] As a result of recent research, it has been demonstrated that endogenously loaded EVs are capable of transporting big proteins and treating neurons. Zhou et al. took MSCs and collected their extracellular vesicles (EVs) in one of their studies. In this study, a novel therapy for cerebral ischemia was studied. It is common knowledge that BDNF possesses effects that are neuroprotective. Because it is unable to pass through the blood-brain barrier, the high-level chemical molecule known as BDNF degrades very quickly. Given this, clinical translation necessitates the implementation of an efficient BDNF delivery mechanism. MSCs have the ability to regulate the immune system. The particular characteristics of the EVs may have significant effects in the afflicted areas. Zhou et al. discovered that intranasally administered extracellular vectors (EVs) targeted the peri-infarct region of an ischemic stroke mouse model due to the attraction of chemokines and the permeability of the brain. Increases in neurogenesis, angiogenesis, synaptic plasticity, and fiber preservation were observed as a result of focused administration.[18] miRNAs and other biological molecules have the ability to deactivate the pathways that are responsible for neuronal death to occur. The researchers Shen et al. evaluated the potential of miR-410 to treat hypoxic/ischemic brain injury (HIBD) by inserting it into bone marrow-derived mesenchymal stem cells (MSCs). The researchers discovered that separated EVs were able to treat primary neurons in vitro by targeting histone deacetylase and deactivating the WNT pathway, which is responsible for the death of neuronal cells. When EVs loaded with mi-R410 were administered to HIBD mice, cognitive performance was likewise improved.[63]
EVs that have been loaded with genetically modified DNA have been the subject of extensive research because of their potential to improve treatment for a wide variety of common diseases. Among these are issues pertaining to the respiratory system, the kidneys, the musculoskeletal system, dermatology, inflammation, bacteria, and metabolism. [Table 2] provides a summary of these investigations.[21], [17], [18], [22], [23], [24], [25], [26], [27], [92], [28], [29], [30], [31], [32], [33], [34], [35], [36], [37], [38], [39], [40], [41], [42], [43], [93], [94], [95], [96], [97], [98], [99], [100], [44], [45], [85], [83], [52], [63], [79], [46], [53], [61], [86], [87], [88], [89], [47], [60], [90], [55], [91], [69], [81], [84] which can be found here.MSCs are preferred for EV generation due to their regenerative, anti-inflammatory, and immunoregulatory capabilities. Inflammatory illnesses include fibrosis, IBD, and osteoarthritis benefit from these features. [17], [81], [72] Several teams have added biologic drugs to MSC-derived EVs to boost their therapeutic potential. Researchers found that EVs supplemented with glial cell line-derived neurotrophic factor (GDNF) increased MSCs' renal fibrosis-reduction effects. They did this by increasing angiogenesis and reducing fibrosis. [17] EVs supplied with IL-27 from parental MSCs via lentivirus transduction improve intestinal epithelial barrier integrity and diminish neutrophil activity in IBD mice models [81]. MSC-derived EVs are angiogenic and aid wound healing. EVs can be loaded with molecules to improve vascularization.[77], [78]
Since macrophages regulate immunity and inflammation, they have also been used to generate EVs. Membrane proteins on macrophage-derived EVs can lead them to inflammation. [101] Tang et al. found that EVs loaded with IL-10 from genetically engineered macrophages targeted the injured kidney in a mouse model of ischemia acute renal injury. This tailored administration reduced renal tubular damage, inflammation, and chronic kidney disease progression. [20] Additional study has examined using macrophage-derived EVs to treat inflammatory disorders. According to Guo et al., miR370-3p-loaded EVs showed promising outcomes in decreasing inflammation and boosting cell survival and proliferation in an IL-1β-induced chondrocyte model Osteoarthritis treatment may be possible with this discovery. [71] Last, EVs can convey the CRISPR/Cas9 technology to cure genetic diseases. This method uses a Cas9 protein and a guide RNA to edit DNA accurately to correct genetic mutations. The lack of secure, precise, and efficient delivery techniques makes EV use similar to CRISPR/Cas9 in clinical contexts. Recent studies show that EVs may carry and administer CRISPR/Cas9 DNA, RNA, and RNP for gene editing in vitro and in vivo. [102] RNPs are favored for endogenous loading because they avoid transcription and translation. This has made gene editing faster, more efficient, and less prone to have off-target consequences. [103] As indicated, scientists are targeting modified RNPs and EVs to increase RNPs.[37] In a recent work, Yao et al. examined the in vivo activity of genetically altered loaded EVs that delivered RNPs targeting exon 53 of the DMD gene. Human DMD gene mutation in exon 53 mice received RNP-loaded EVs, which decreased skeletal muscle dystrophin synthesis. The study found that RNP-loaded EVs can change genes and increase dystrophin expression. [76] This novel method shows how EVs can transport the CRISPR/Cas9 system for precise DNA editing, especially for hereditary diseases.
Conclusion
There are many reasons EVs are good for transporting large macromolecules like proteins and genetic material. Several of these compounds have been loaded onto EVs and delivered to particular cells, demonstrating their therapeutic potential in lab and live tests. The preclinical investigations have not been tested in humans. Therefore, certain obstacles must be addressed to facilitate drug delivery clinical development. The variability of EV content, the lack of standardized production and isolation procedures, and the difficulties in scaling up production stand out. This review highlights genetic engineering as a strong tool for turning parental cells into efficient manufacturers producing stable and consistent extracellular vesicles. EVs can also be loaded with therapeutic proteins and nucleic acids using this method. This loading method enhances EV batch uniformity, surface structure, and time and effort over the exogenous method. In addition, parental gene engineering-based loading techniques have opened up new ways to solve biological therapy difficulties. Overall, the findings examined will influence the development of new medications. While many present bioengineering technologies concentrate single-step procedures, it is obvious that synergistic cargo loading and delivery approaches will substantially increase EV functionalization in the future. EV biology, notably biogenesis, systemic dispersion, and cell transit, must be understood. This understanding is important to maximize their therapeutic potential as future nanodelivery technologies.
Source of Funding
None.
Conflict of Funding
None.
References
- Lu M, HX, Zheng A, YH, XL. Overcoming pharmaceutical bottlenecks for nucleic acid drug development. Acc Chem Res. 2023;56(3):224-36. [Google Scholar]
- Wang C, Zhang Y. Lipid nanoparticle-mRNA formulations for therapeutic applications. Acc Chem Res. 2021;54(23):4283-93. [Google Scholar]
- Liu J, CZ, XZ, Yan J, Zeng C, Talebian F. Intratumoral delivery of IL-12 and IL-27 mRNA using lipid nanoparticles for cancer immunotherapy. J Control Release. 2022;345:306-13. [Google Scholar]
- Zeng Y. Biological features of extracellular vesicles and challenges. Front Cell Dev Biol. 2022;10. [Google Scholar]
- Moloudizargari M. The therapeutic triad of extracel- lular vesicles: as drug targets, as drugs, and as drug carriers. Biochem Pharmacol. 2021;192. [Google Scholar]
- Popowski K. Inhalable exosomes outperform liposomes as mRNA and protein drug carriers to the lung. Extracell Vesicle. 2022;1. [Google Scholar] [Crossref]
- Wang B. RNA and protein delivery by cell-secreted and bioengineered extracellular vesicles. Adv. Healthc. Mater. 2021;11(5). [Google Scholar]
- Hettich B. Encapsulation of hydrophilic com- pounds in small extracellular vesicles: loading capacity and impact on vesicle functions. Adv Healthc Mater. 2022;11(5). [Google Scholar]
- Reshke R, Taylor J, Savard A, Guo H, Rhym L, PK. Reduction of the therapeutic dose of si- lencing RNA by packaging it in extracellular vesicles via a pre- microRNA backbone. Nat Biomed Eng. 2020;4(1):52-68. [Google Scholar]
- Dumontel B. Nanotechnological engineering of extracellular vesicles for the development of actively targeted hybrid nanodevices. Cell Biosci. 2022;12(1). [Google Scholar] [Crossref]
- Colja S. Sonication is a suitable method for loading nanobody into glioblastoma small extracellular vesicles. Heliyon. 2023;9(5). [Google Scholar]
- Wiklander O. Advances in therapeutic applications of extracellular vesicles. Sci Transl Med. 2019;11(492). [Google Scholar]
- Wu P. Extracellular vesicles: a bright star of nano- medicine. Biomaterials. 2021;269. [Google Scholar]
- Ma H. Extracellular vesicle-mediated delivery of miR-127-3p inhibits the proliferation and invasion of choriocar- cinoma cells by targeting ITGA6. Exp Cell Res. 2022;414(2). [Google Scholar]
- Yu X. Extracellular vesicle-mediated delivery of circDYM alleviates CUS-induced depressive-like behaviours. J Extracell Vesicle. 2022;11(1). [Google Scholar]
- Cecchin R, Troyer Z, Witwer K, Morris K. Extracellular vesicles: the next generation in gene therapy delivery. Mol Ther. 2023;31(5):1225-30. [Google Scholar]
- Chen L, YW, Shulin L, BZ, Zhang X, Wang F. Exosomes derived from GDNF-modified human adipose mesenchymal stem cells ameliorate peritubular capillary loss in tubulointerstitial fibrosis by activating the SIRT1/ eNOS signaling pathway. Theranostics. 2020;10(20):9425-42. [Google Scholar]
- Zhou X. Intranasal delivery of BDNF-loaded smallextracellular vesicles for cerebral ischemia therapy. J Control Release. 2023;357:1-19. [Google Scholar]
- Rossowska J. Antitumor potential of extracellular vesicles released by genetically modified murine colon carci- noma cells with overexpression of interleukin-12 and shRNA for TGF-β1. Front Immunol. 2019;10. [Google Scholar] [Crossref]
- Tang T. Extracellular vesicle-encapsulated IL-10 as novel nanotherapeutics against ischemic AKI. Sci Adv. 2020;6(33). [Google Scholar]
- Salazar-Puerta A. Engineered extracellular vesi- cles derived from dermal fibroblasts attenuate inflammation in a murine model of acute lung injury. Adv Mater. 2023;35(28). [Google Scholar]
- Lu W. Engineered NF-κB siRNA-encapsulating exosomes as a modality for therapy of skin lesions. Front Immunol. 2023;14. [Google Scholar]
- Teng F, Fussenegger M. Shedding light on extracel- lular vesicle biogenesis and bioengineering. Adv Sci (Weinh). 2020;8(1). [Google Scholar]
- Leidal A, Debnath J. Unraveling the mechanisms that specify molecules for secretion in extracellular vesicles. Methods. 2020;177:15-26. [Google Scholar]
- Zheng W, Rädler J, Sork H, Niu Z, Roudi S, Bost J. Identification of scaffold proteins for im- proved endogenous engineering of extracellular vesicles. Nat Commun. 2023;14. [Google Scholar]
- Rädler J, Gupta D, Zickler A, SEA. Exploiting the biogenesis of extracellular vesicles for bioengineering and therapeutic cargo loading. Mol Ther. 2023;31(5):1231-50. [Google Scholar]
- Sterzenbach U. Engineered exosomes as vehiclesfor biologically active proteins. Mol Ther. 2017;25(6):1269-78. [Google Scholar]
- Zuppone S, Zarovni N, Vago R. The cell type dependent sorting of CD9- and CD81 to extracellular vesicles can be exploited to convey tumor sensitive cargo to target cells. Drug Deliv. 2023;30(1). [Google Scholar]
- Corso G. Systematic characterization of extracellular vesicle sorting domains and quantification at the single molecule - single vesicle level by fluorescence correlation spectroscopy and single particle imaging. J Extracell Vesicles. 2019;8(1). [Google Scholar]
- Silva A, EI, Gunnarsson A, AD, Daaboul G, BP. Quantification of protein cargo loading into engineered extracellular vesicles at single-vesicle and single-molecule resolution. J Extracell Vesicles. 2021;10(10). [Google Scholar]
- Do M, Levy D, Brown A, GM, Lu B. Targeted delivery of lysosomal enzymes to the endocytic compartment in human cells using engineered extracellular vesicles. Sci Rep. 2019;9(1):17274-85 . [Google Scholar]
- Levy D. Orchestrating extracellular vesicle with dual reporters for imaging and capturing in mammalian cell cul- ture. Front Mol Biosci. 2021;8. [Google Scholar]
- Zhang X, QX, Zi Z, Liu Z, CW, Crisman L. Programmable extracellular vesicles for macromolecule delivery and genome modifications. Dev Cell. 2020;55(6):784-801. [Google Scholar]
- Somiya M, Kuroda S. Engineering of extracellularvesicles for small molecule-regulated cargo loading and cytoplas- mic delivery of bioactive proteins. Mol Pharm. 2022;19(7):2495-505. [Google Scholar]
- Ilahibaks N. TOP-EVs: technology of protein delivery through extracellular vesicles is a versatile platform for intracellular protein delivery. J Control Rel. 2023;355:579-92. [Google Scholar]
- Bui S, Dancourt J, Lavieu G. Virus-free method to control and enhance. ACS Appl Bio Mater. 2023;6(3):1081-91. [Google Scholar]
- . extracellular vesicle cargo loading and delivery. ACS Appl. Bio Mater. ;6:1081-1091. [Google Scholar]
- Dooley K. A versatile platform for generating engineered extracellular vesicles with defined therapeutic prop- erties. Mol. Ther. 2021;29:1729-1743. [Google Scholar]
- El Amri M. MARCKS and MARCKS-like proteins in development and regeneration. J. Biomed. Sci. 2018;25:43-43. [Google Scholar]
- Liu T. EVmiRNA: a database of miRNA profiling in extracellular vesicles. Nucleic Acids Res. 2019;47:89-93. [Google Scholar]
- Garcia-Martin R. MicroRNA sequence codes for small extracellular vesicle release and cellular retention. Nature. 2022;601:446-451. [Google Scholar]
- Dixson AC. Context-specific regulation of extracellular vesicle biogenesis and cargo selection. Nat. Rev. Mol. Cell Biol. 2023;24:454-476. [Google Scholar]
- Ramos D. Extracellular vesicle-based delivery of silencing sequences for the treatment of Machado-Joseph disease/spinocerebellar ataxia type 3. Mol Ther. 2023;31(5):1275-92. [Google Scholar]
- Mi B, Chen L, Xiong Y, YY. Osteoblast/osteoclast and immune cocktail therapy of an exosome/drug delivery multifunctional hydrogel accelerates fracture repair. ACS Nano. 2022;16(1):771-82. [Google Scholar]
- Es-Haghi M. Construction of fusion protein for enhanced small RNA loading to extracellular vesicles. Genes. 2023;14(2). [Google Scholar]
- Zhu C. Tumor-derived extracellular vesicles inhibit HGF/c-Met and EGF/EGFR pathways to accelerate the radio- sensitivity of nasopharyngeal carcinoma cells via microRNA- 142-5p delivery. Cell Death Discov. 2022;8(1). [Google Scholar]
- Guo Z, YZ, Xu W, Zhang X, Jiang J. Engineered exosome-mediated delivery of circDIDO1 inhibits gastric cancer progression via regulation of MiR-1307-3p/SOCS2 axis. J Transl Med. 2022;20(1). [Google Scholar]
- Liu Y, RG, Yan J, Zhu Y, Sun S, Qu Y. Mesenchymal stem cell-derived extracellular vesicle-shuttled microRNA-302d-3p represses inflammation and cardiac remodeling following acute myocardial infarction. J Cardiovasc Transl Res. 2022;15(4):754-71. [Google Scholar]
- Xiao Z. EVs delivery of miR-1915-3p improves the chemotherapeutic efficacy of oxaliplatin in colorectal cancer. Cancer Chemother Pharmacol. 2021;88(6):1021-31. [Google Scholar]
- Kimura Y, Ohzawa H, Miyato H, Kaneko Y, Kuchimaru T, Takahashi R. Intraperitoneal transfer of microRNA- 29b-containing small extracellular vesicles can suppress peritoneal metastases of gastric cancer. Cancer Sci. 2023;114(7):2939-50. [Google Scholar]
- Dancourt J, Piovesana E, Lavieu G. Efficient cell death mediated by bioen- gineered killer extracellular vesicles. Sci Rep. 2023;13(1). [Google Scholar]
- El-Hage N, MH, Zhao U, Rodriguez M, Wu Z, Liu M. Extracellular vesicles released by genetically modified macrophages activate autophagy and pro- duce potent neuroprotection in mouse model of lysosomal storage disorder. Batten Dis Cells. 2023;12(11). [Google Scholar]
- Chen J. Delivery of miR-130a-3p through adipose- derived stem cell-secreted EVs protects against diabetic peripheral neuropathy via DNMT1/NRF2/HIF1α/ACTA1 axis. Mol Neurobiol. 2023;60(7):3678-94. [Google Scholar]
- Zhao J. MiR-26a-5p from HucMSC-derived extracellular vesicles inhibits epithelial mesenchymal transition by targeting Adam17 in silica-induced lung fibrosis. Ecotoxicol Environ Saf. 2023;257. [Google Scholar] [Crossref]
- Zhang J, Su R, Wang Y, Wang H, SL. Protective effect of small extracellular vesi- cles (EVs) derived from ACE2-modified human umbilical cord mesenchymal stem cells against renal ischemia-reperfusion injury. Nephrology (Carlton). 2024;29(1):5-17. [Google Scholar]
- Yang Y, Wang J, Zhang Y, Hu X. Exosomes derived from mesenchymal stem cells ameliorate renal fibrosis via delivery of miR-186-5p. Hum Cell. 2022;35(12):83-97. [Google Scholar]
- Saint-Pol J. Targeting and crossing the blood- brain barrier with extracellular vesicles. Cells. 2020;9(4). [Google Scholar]
- Zheng H. Exosome-encapsulated miR-26a atten- uates aldosterone-induced tubulointerstitial fibrosis by inhibiting the CTGF/SMAD3 signaling pathway. Int J Mol Med. 2023;51(2). [Google Scholar]
- Tang T, Lv LL, Wang B, Cao J, YF, Li ZL. Employing macrophage-derived microvesicle for kidney-targeted delivery of dexamethasone: an efficient therapeutic strategy against renal inflammation andfibrosis. Theranostics. 2019;9(16):4740-55. [Google Scholar]
- Yang Z, Ji P, Li Z, Zhang R, Wei M, Yang Y. Improved extracellular vesicle-based mRNA delivery for familial hypercholesterolemia treatment. Theranostics. 2023;13(10):3467-79. [Google Scholar]
- Liu Y, YZ, Si HB, Tang L, Xie HQ, BS. Exosomes derived from human urine- derived stem cells overexpressing miR-140-5p alleviate knee osteoarthritis through downregulation of VEGFA in a rat model. Am J Sports Med. 2022;50(4):1088-105. [Google Scholar]
- Yu X. Bone marrow mesenchymal stem cell derived extracellular vesicles carrying circ_0050205 attenuate intervertebral disc degeneration. Oxid Med Cell Longev. 2022. [Google Scholar] [Crossref]
- Tang S. Non-viral reprogramming of human nucleus pulposus cells with FOXF1 via extracellular vesicle delivery: an in vitro and in vivo study. Eur Cell Mater. 2021;41:90-107. [Google Scholar]
- Liang Y, Iqbal Z, Wang J, Xu L, Xu X, Ouyang K. Cell-derived extracellular vesicles for CRISPR/Cas9 delivery: engineering strategies for cargo packaging and loading. Biomater Sci. 2022;10:4095-106. [Google Scholar]
- Lin Z, Ma Y, Zhu X, Dai S, Sun W, Li W. Potential predictive and therapeutic applications of small extracellular vesicles-derived circPARD3B in osteoarthritis. Front Pharmacol. 2022;13. [Google Scholar]
- Li Q, Hu W, Huang Q, Yang J, Li B, Ma K. MiR146a-loaded engineered exosomes re- leased from silk fibroin patch promote diabetic wound healing by targeting IRAK1. Signal Transduct Target Ther. 2023;8(1). [Google Scholar]
- Shen M. Neuroprotection of bone marrow-derived mesenchymal stem cell-derived extracellular vesicle-enclosed miR-410 correlates with HDAC4 knockdown in hypoxic- ischemic brain damage. Neurochem Res. 2022;47(10):3150-66. [Google Scholar]
- Yang Z. Hydrogel armed with Bmp2 mRNA-enriched exosomes enhances bone regeneration. J. Nanobiotechnol. 2023;21:119-119. [Google Scholar]
- Yu L, Sui B, Zhang X, Liu J, Hao X, Hao X. miR-92a-1-5p enriched prostate cancer ex- tracellular vesicles regulate osteoclast function via MAPK1 and FoxO1. J Exp Clin Cancer Res. 2023;42(1). [Google Scholar]
- Y. Intradermally delivered mRNA-encapsulating extracellular vesicles for collagen-replacement therapy. Nat. Biomed. Eng. 2023;7:887-900. [Google Scholar]
- Xu M. Recent advancements in the loading and modification of therapeutic exosomes. Front Bioeng Biotechnol. 2020;8. [Google Scholar]
- Nie M. Bioadhesive microcarriers encapsulated with IL-27 high expressive MSC extracellular vesicles for inflammatory bowel disease treatment. Adv. Sci. (Weinh). 2023;10:2303349-2303349. [Google Scholar]
- Gao W, Zhang Y, Yuan I, Huang F, Wang YS. Long non-coding RNA H19-overexpressing exosomes ameliorate UVB-induced photoaging by upregulating SIRT1 via sponging miR-138. Photochem Photobiol. 2023;99(6):1456-7. [Google Scholar]
- Blavier L. The capture of extracellular vesiclesendogenously released by xenotransplanted tumours induces an inflammatory reaction in the premetastatic niche. J Extracell Vesicles. 2023;12(5). [Google Scholar]
- Moradi-Chaleshtori M. Overexpression of pigment epithelium-derived factor in breast cancer cell-derived exosomes in- duces M1 polarization in macrophages. Immunol Lett. 2022;248:31-6. [Google Scholar]
- Pan Z, Chen Q, Ding H, Li H. MicroRNA-342-3p loaded by human umbilical cord mesenchymal stem cells-derived exosomes attenuates deep vein thrombosis by downregulating EDNRA. J Thromb Thrombolysis. 2022;54(3):411-9. [Google Scholar]
- Wang Q, Dong Y, Wang H. MicroRNA-19b-3p-containing extracellular vesicles derived from macrophages promote the development of atherosclerosis by targeting JAZF1. J Cell Mol Med. 2022;26(1). [Google Scholar]
- Chen C, YL, QW, Cai N, Wu L, Yan X. Single-particle assessment of six different drug-loading strategies for incorporating doxorubicin into small extracellular vesicles. Anal Bioanal Chem. 2023;415(7):1287-98. [Google Scholar]
- Shrivastava S, Ray R, Holguin L, Echavarria L. Exosome-mediated stable epige- netic repression of HIV-1. Nat Commun. 2021;12. [Google Scholar]
- Mao S, Zhao J, Zhang J. MiR-183-5p overexpression in bone mesenchymal stem cell-derived exosomes protects against myocardial ischemia/reperfusion injury by targeting FOXO1. Immunobiology. 2022;227(3). [Google Scholar]
- Wasylewska K. Induced pluripotent stem cell-derived extracellular vesicles enriched with miR-126 induce proangiogenic properties and promote repair of ischemic tissue. Fase J. 2024;38(2). [Google Scholar]
- Lange M, Lyddon T, MJ. Diphtheria toxin A-resistant cell lines enable robust production and evaluation of DTA-encoding lentiviruses. Sci Rep. 2019;9(1). [Google Scholar]
- Xuan B, Wang Y. Exosome-transmitted miR-506- 3p inhibits colorectal cancer cell malignancy via regulating GSTP1. Appl Biochem Biotec. 2023;195(3):2015-27. [Google Scholar]
- Chen J, Ding C, Yang X, Zhao J. BMSCs-derived exosomal MiR-126-3pinhibits the viability of NSCLC cells by targeting PTPN9. J Buon. 2021;26(5):1832-41. [Google Scholar]
- Biswas J. Zipcode binding protein 1 (ZBP1; IGF2BP1): a model for sequence-specific RNA regulation. Cold Spring Harb. Symp Quant Biol. 2019;84:1-10. [Google Scholar]
- Joshi B, Youssef S, Bron R, AdB, Kampinga H, Zuhorn I. DNAJB6b-enriched small extracellular vesicles decrease polyglutamine aggregation in in vitro and in vivo models of Huntington disease. iScience. 2021;24(11). [Google Scholar]
- Luo T. Transcriptomic features in a single extracel- lular vesicle via single-cell RNA sequencing. Small Methods. 2022;6(11). [Google Scholar]
- Jiang X. Extracellular vesicles derived from human ESC-MSCs target macrophage and promote anti-inflammation process, angiogenesis, and functional recovery in ACS-induced severe skeletal muscle injury. Stem Cell Res Ther. 2023;14. [Google Scholar]
- Buzas E. The roles of extracellular vesicles in the immune system. Nat Rev Immunol. 2023;23:236-50. [Google Scholar]
- Xing Y, Zhang F, Ji P, Yin H, Yin H, Yang A. Efficient delivery of GSDMD-N mRNA by engineered extracellular vesicles induces pyroptosis for enhanced immunotherapy. Small. 2023;19(20). [Google Scholar]
- Pang Y. Engineered multitargeting exosomes carrying miR-323a-3p for CRC therapy. Int J Biol Macromol. 2023;247. [Google Scholar]
- Cheng Y, Lu X, FL, Chen Z, Zhang Y, Han Q. NDFIP1 limits cellular TAZ accumulation via exosomal sorting to inhibit NSCLC proliferation. Protein Cell. 2023;14(2):123-36. [Google Scholar]
- Hu Z, Chen G, Zhao Y. Exosome-derived circCCAR1 promotes CD8+ T-cell dysfunction and anti-PD1 resistance in hepatocellu- lar carcinoma. Mol Cancer. 2023;22(1). [Google Scholar]
- Kim B. SYNCRIP controls miR-137 and striatal learning in animal models of methamphetamine abstinence. Acta Pharm Sin B. 2022;12(8):3281-97. [Google Scholar]
- Shirazi S, Huang C, Kang M, Lu Y, Ravindran S, Cooper L. The importance of cellular and exosomal miRNAs in mesenchymal stem cell osteoblastic differentiation. Sci Rep. 2021;11(1). [Google Scholar]
- Barman B, Sung B. VAP-A and its binding partner CERT drive biogenesis of RNA-containing extracellular vesicles at ER membrane contact sites. Dev Cell. 2022;57(8):974-94. [Google Scholar]
- Liu X. Selective sorting of microRNAs into exosomes by phase-separated YBX1 condensates. Elife. 2021;10. [Google Scholar]
- Lin F, Zeng Z. YBX-1 mediated sorting of miR-133 into hypoxia/reoxygenation-induced EPC-derived exosomes to in- crease fibroblast angiogenesis and MEndoT. Stem Cell Res Ther. 2019;10:263-8. [Google Scholar]
- Li C, Wang W, YS. Selective sorting and secretion of hY4 RNA fragments into extracellular vesicles mediated by methylated YBX1 to promote lung cancer progression. J Exp Clin Cancer Res. 2022;41(1). [Google Scholar]
- Wozniak A. The RNA binding protein FMR1 controls selective exosomal miRNA cargo loading during inflammation. J Cell Biol. 2020;219. [Google Scholar]
- Born L, Chang K, Shoureshi P, Lay F, Bengali S, Hsu ATW. HOTAIR-loaded mesenchymal stem/ stromal cell extracellular vesicles enhance angiogenesis and wound healing. Adv Healthc Mater. 2022;11(5). [Google Scholar]
- Guo W, Su L, Zhang H, Mi Z. Role of M2 macrophages-derived extra- cellular vesicles in IL-1β-stimulated chondrocyte proliferation and inflammatory responses. Cell Tissue Bank. 2023;24(1):93-107. [Google Scholar]
- Lu Y. CRISPR-Cas9 delivery strategies with engineered extracellular vesicles. Mol Ther Nucleic Acids. 2023;34. [Google Scholar] [Crossref]
- Abstract
- EV Nanoprobes Have the Ability to Supply Biomolecules
- Loading Instructions for Endogenous Extracellular Vesicles
- Enhancing EV Biologic Drug Loading Via Engineering
- Methods developed recently to improve EV protein medication loading
- The most recent techniques for enhancing the drug loading of EV nucleic acid
- Treatment using EVs loaded with GMOs.
- Genetically Modified Extracellular Vesicles for Various Diseases
- Transportation vehicles that Have been genetically modified for the purpose of treating cardiovascular disease
- EVs that fight cancer have been genetically engineered
- EVs are fascinating as potential therapies for neurological problems
- Conclusion
- Source of Funding
- Conflict of Funding
- References